Cornus sericea
Mant. Pl. 2: 199. 1771.

Shrubs, to 4 m, flowering at 1 m; rhizomes absent. Stems clustered, branches occasionally arching to the ground and rooting at nodes; bark yellow to red, not corky, loosely verrucose; branchlets bright red, reddish brown, maroon, or green, occasionally green in winter and maroon in summer, appressed-hairy when young; lenticels protruding on 2d year branches, area surrounding them not suffused with purple on older branches; pith white. Leaves: petiole 5–38 mm; blade lanceolate, elliptic, or ovate, 3.5–20 × 1.5–12 cm, base cuneate, apex acute to acuminate, abaxial surface white, hairs appressed except near secondary vein axils, tufts of erect hairs present in axils of secondary veins, adaxial surface green, hairs appressed, sparse; secondary veins 5–7 per side, most arising from proximal 1/2, tertiary veins not prominent. Inflorescences flat-topped, 3–6 cm diam., peduncle 20–40 mm; branches and pedicels green to yellow-green, turning maroon in fruit. Flowers: hypanthium densely appressed-hairy; sepals 0.2–0.6 mm; petals white to cream, 2.5–4 mm. Drupes white, globose or subglobose, 6–10 mm diam.; stone subglobose, laterally compressed, 4–6 × 4–6 × 1.5–3 mm, furrowed laterally, apex rounded. 2n = 22.
Phenology: Flowering May–Jun and Sep–Oct; fruiting Aug–Oct.
Habitat: Wet meadows, thickets, edges of mesic upland forests, fens, marshes, swamps, stream banks, lake shores, river banks.
Elevation: 0–2500 m.
Distribution
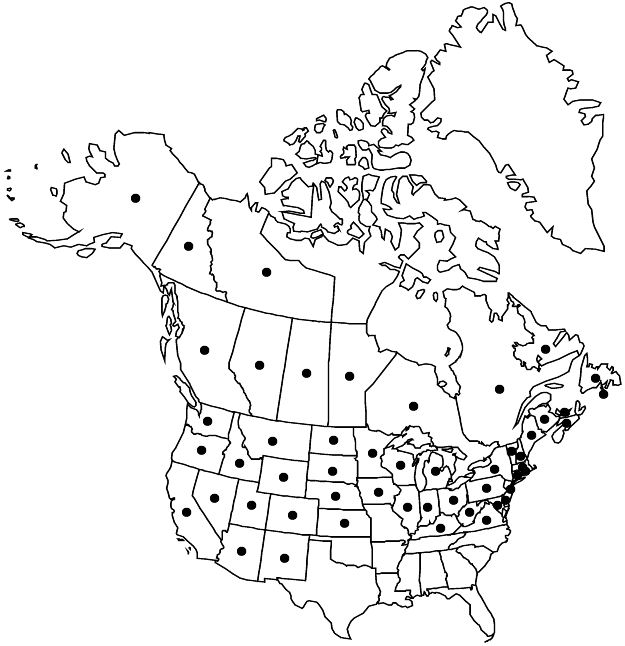
St. Pierre and Miquelon, Alta., B.C., Man., N.B., Nfld. and Labr., N.W.T., N.S., Ont., P.E.I., Que., Sask., Yukon, Alaska, Ariz., Calif., Colo., Conn., Del., Idaho, Ill., Ind., Iowa, Kans., Ky., Maine, Md., Mass., Mich., Minn., Mont., Nebr., Nev., N.H., N.J., N.Mex., N.Y., N.Dak., Ohio, Oreg., Pa., R.I., S.Dak., Utah, Vt., Va., Wash., W.Va., Wis., Wyo., Mexico (Chihuahua, Durango, Nuevo León), introduced w Europe.
Discussion
As the synonymy implies, Cornus sericea has received considerable attention from taxonomists wishing to subdivide the species, presumably in order to make it more comprehensible. Most of the divisions have been based upon indumentum and stone differences, although habit has also been used. Although one of the synonyms and one of the common names imply a stoloniferous habit, the species is not stoloniferous; evidently, branch tips infrequently arching to the ground and rooting at the nodes led to confusion regarding the growth habit. H. W. Rickett (1944b) examined the morphology of the various forms, varieties, and subspecies, and found extensive overlap using fruit shape and indumentum differences. It is not known whether the variation is due to primary differentiation or secondary intergradation, and the complex is treated here as a single species. There is little doubt that the European species C. alba Linnaeus is closely related to C. sericea and should be included in any future studies of this species complex.
The name Cornus stolonifera has sometimes been applied to C. sericea (for example, H. W. Rickett 1944b) because the description by Linnaeus of the latter could apply to several currently recognized species. F. R. Fosberg (1942) lectotypified C. sericea, establishing that it applies to this species.
Cornus sericea is commonly planted as an ornamental and occasionally escapes; plants in suburban areas and in highly acidic soils are suspected as non-natural occurrences. Putative hybrids between C. sericea and C. rugosa have been called C. ×slavinii Rehder, and are reported from Maine, New York, Ontario, Prince Edward Island, and Wisconsin.
Selected References
None.