Spiranthes cernua
De Orchid. Eur., 37. 1817.

Plants 10–50 cm. Roots few, slender to tuberously thickened, horizontally spreading to descending, mostly to 1 cm diam. Leaves persisting through anthesis or fugaceous, basal, sometimes on proximal portion of stem, ascending to spreading (flaccidly so because of membranaceous blades with thickened midribs); petiole usually slender, less than 6 mm wide; blade linear-lanceolate to linear-oblanceolate, oblanceolate, obovate, or elliptic, to 26 × 2 cm. Inflorescences: spikes very tightly to rather loosely spiraled, 3–4 flowers per cycle of spiral, rarely in loose spiral with 5 or more flowers per cycle; rachis moderately to densely pubescent, some trichomes capitate, glands obviously stalked. Flowers white to ochroleucous, creamy, or ivory, occasionally green or rarely yellowish, nodding only at base of perianth or occasionally ascending, urceolate to gaping, base usually appearing only slightly inflated, claw of lip at low (less than 30°) angle to lamina, or occasionally not opening; sepals distinct to base, 6–12 mm; lateral sepals appressed to spreading, apices incurved or sometimes straight, sometimes arching above flower; petals linear-lanceolate, 6–12 mm, apex acute to obtuse, occasionally broad and liplike; lip often yellow centrally, ovate to oblong in general outline, usually constricted at middle, 6–10.5 × 2–7.5 mm, base ± prominently dilated, suborbiculate, orbiculate, or linear, margins crenulate, lacerate or sometimes entire, apex acute to rounded distally, glabrous; veins several, branches parallel; basal calli usually incurved, prominent to conic, small, mostly 0.5–1.2 mm, or sometimes lip undifferentiated from petals; viscidia linear to linear-lanceolate; ovary 3–14 mm. Seeds polyembryonic, often also monoembryonic, rarely wholly monoembryonic; typically showing irregularities such as embryos developing through or outside testa or naked embryos mixed with whole seed. 2n = 45, 60.
Phenology: Flowering late Aug–Nov.
Habitat: Wet to dry open sites in fens, marshes, meadows, swales, dunes, prairies, open woodlands, riverbanks, shores, ditches, roadsides, old fields, cemeteries, lawns
Elevation: 0–1800 m
Distribution
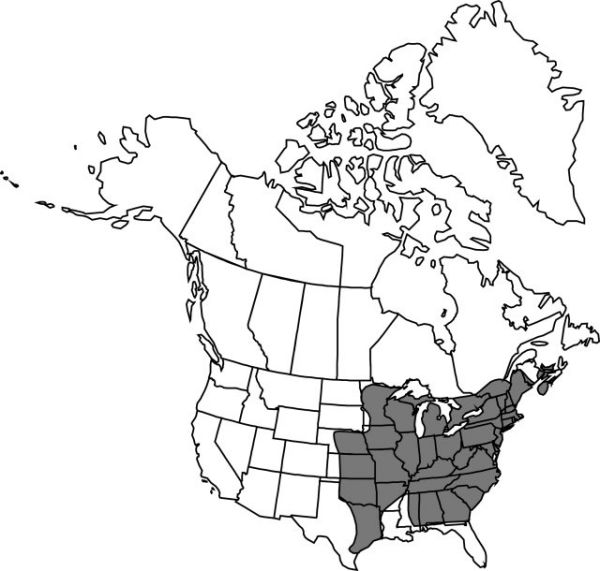
N.B., N.S., Ont., P.E.I., Que., Ala., Ark., Conn., Ga., Ill., Ind., Iowa, Kans., Ky., La., Maine, Md., Mass., Mich., Minn., Miss., Mo., Nebr., N.H., N.J., N.Y., N.C., Ohio, Okla., Pa., R.I., S.C., Tenn., Tex., Vt., Va., W.Va., Wis.
Discussion
Spiranthes cernua is a facultatively agamospermic polyploid compilospecies in which unidirectional gene flow from related diploids generates a wide range of novel forms and races. Chromosomal segregation has given rise to some forms that very nearly approximate the diploid donor, and meiotic irregularities produce peloria and other abnormal conditions. Apomixis then permits reproduction in pure lines, some of which are so distinctive as to appear to be species. Determination of this and related species is consequently often challenging.
Adventitious embryony in this species commonly gives rise to polyembryonic seeds and seeds exhibiting irregularities such as embryos erupting through or developing outside the testa and naked embryos lacking testae. Often a capsule contains a mixture of polyembryonic and monoembryonic seeds. Rarely all or most seed is monoembryonic, yet irregularities can still be seen.
Polyembryony is also characteristic of Spiranthes casei and S. parksii, species that are probably derivatives of S. cernua. For the most part, at least, related diploids and all other species are strictly sexual and characterized by wholly monoembryonic seed. Some indication of apomixis and polyembryony in S. magnicamporum, S. ochroleuca, and S. odorata in the northeastern portion of the range of the complex has been reported (P. M. Catling 1981, 1982), but the difficulty in distinguishing between diploid donors and tetraploid chromosomal segregates makes interpretation difficult (C. J. Sheviak 1982, 1991). In particular, the range of S. odorata northward along the Atlantic Coast is uncertain. Plants that appear typical of S. odorata in all respects but with polyembryonic seed occur from Virginia northward and quickly replace monoembryonic plants toward the north. Such plants from as far south as Virginia, however, have proven to be tetraploids and furthermore to assume morphology typical of S. cernua when grown away from the Coast.
Gene flow from Spiranthes magnicamporum generates cytogenetically based peloria, a common element in portions of the prairie region. Most commonly differentiation of the lip is suppressed, but in some forms the petals are elaborated and approach the lip in form. Additionally, buds may fail to open and a functional column may be lacking, yet seed is produced apomictically. See the discussion under 16. S. parksii.