Dioscorea villosa
Sp. Pl. 2: 1032. 1753.

Plants rhizomatous; rhizomes brownish, unbranched or highly branched, linear to irregularly contorted, 0.5–1.5 cm or more diam., nodes not articulate. Stems longitudinally grooved or sometimes narrowly winged, usually terete in cross section, 1–7 m, ± rigid proximally, or flexible, glabrous or rarely with sparse pubescence, wings when present less than 1 mm and stems polygonal. Leaves alternate, subopposite, subverticillate, or in verticels of 3–7 proximally, due to suppression of proximal internodes (esp. in woodland understory), always alternate distally, 3–13 × 2–13 cm, ca. as long as wide; petiole ridged or narrowly winged, 3–14 cm, glabrous or puberulent at pulvinus, base not clasping; blade green to ± glaucous, (7–)9–11-veined, ovate-cordate, abaxial surface sometimes ± glandular, or sparsely or sometimes densely pubescent to glabrous, base with sinus rounded, acute, or ± truncate basally, margins entire or repand, apex acute to acuminate, occasionally mucronate. Staminate inflorescences solitary in leaf axils, rarely terminal, spicate or branched; cymes sessile, bearing 1–3 sessile flowers, braceolate, internodes between cymes 1–8 mm, bracteoles ca. 1 mm; rachis 2–30 cm, secondary axes to 15 cm, robust plants occasionally branched to third order, axes subtended by linear-lanceolate bracts 1–3 mm. Pistillate inflorescences solitary, 4–18-flowered, 4–20 cm, internodes 6–12 mm. Staminate flowers: perianth greenish white, appearing darker in some specimens due to presence of irregularly distributed tannin crystals, rotate-campanulate to funnelform, 1–2(–3) mm diam.; tepals ± glandular, ovate-elliptic, margins hyaline, apex rounded or acute; stamens in 2 subequal whorls, erect; anthers ca. ½ length of filaments, thecae distinct, widely spreading. Pistillate flowers: perianth greenish white, rotate-campanulate, 2–4 mm wide; tepals as in staminate flowers; staminodes 6, differentiated into anthers and filaments, less than 1/2 length of fertile stamens. Capsules greenish gold, ovoid to obovoid to obreniform, 1–3 × 1–3.5 cm, varying continously in size, occasionally ± glaucous. Seeds generally 2 per locule, rarely 1, 5–18 mm. 2n = 20, 36, 54, 60.
Phenology: Flowering mid spring–summer; fruiting late summer.
Habitat: Borders of bogs, swamps, marshes, river and lake margins, creek bottoms, sandy or rocky soils, moist or dry woods, hammocks, thickets, limestone or talus slopes, roadsides
Elevation: 0–1500 m
Distribution
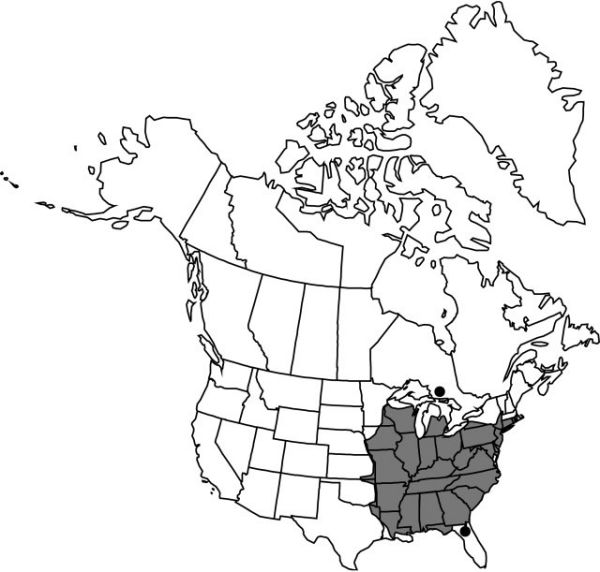
Ont., Ala., Ark., Conn., Del., D.C., Fla., Ga., Ill., Ind., Iowa, Kans., Ky., La., Md., Mass., Mich., Minn., Miss., Mo., Nebr., N.J., N.Y., N.C., Ohio, Okla., Pa., R.I., S.C., Tenn., Tex., Vt., Va., W.Va., Wis.
Discussion
Dioscorea villosa is a highly polymorphic species, exhibiting complex patterns of variation across its geographic range. Characters that have been used previously to delineate taxonomic boundaries within this complex—pubescence, glaucousness, rhizome thickness and shape, length of internodes within the inflorescence, arrangement of proximal leaves, geometry of the stem, and fruit and seed size/shape—fail when individuals from all parts of the range and specimens representing both apical and basal portions of single stems can be examined. At its morphological extremes, D. villosa comprises 1) small vines with tightly congested inflorescences, winged stems, and variously pubescent leaves, occurring in bogs and branch swamps; and 2) robust plants, rigid at the base, the proximal leaves verticillate with large, glaucous blades, from the axils of which arise lax spikes or panicles, inhabiting rocky, upland woods and steep talus slopes. As one ascends from the Atlantic Coastal Plain through the Appalachians, continuing westward to the Great Lakes region, south to the Ozarks, and east to the branch swamps of Georgia, particular morphologies are associated with particular ecological conditions, independent of geography. As well as the morphological extremes, every intermediate condition of leaf, stem, and inflorescence architecture can be found, in all combinations, and variation may be encountered even within individual plants. What sort of genetic structure underlies these patterns of morphological diversity remains an open question. That there is a significant degree of genetic variability within the complex is evident from the chromosome counts thus far reported. Further research is needed to shed light on patterns of gene flow in the complex, and garden studies would be instructive as to the limits of individual plasticity. At present, I can find no natural gaps in the variation between the plants that have been called (albeit ambiguously; see H. H. Bartlett 1910) D. villosa and those called D. quaternata, and therefore I am treating the complex as a single species.
Selected References
None.