Lepidium draba
Sp. Pl. 2: 645. 1753.

Perennials; (rhizomatous); hirsute or glabrate. Stems often simple from base, erect or decumbent basally, branched (several) distally, (0.8–)2–6.5(–9) dm. Basal leaves (early withered); not rosulate; petiole 1–4 cm; blade obovate, spatulate, or ovate, (1.5–)3–10(–15) cm × 10–40 mm, margins sinuate to dentate or entire. Cauline leaves sessile; blade ovate, elliptic, oblong, or lanceolate, oblanceolate, or obovate, (1–)3–9(–15) cm × (5–)10–20(–50) mm, base sagittate-amplexicaul or auriculate, margins dentate or entire, (surfaces pubescent or glabrous). Racemes (corymbose panicles), slightly or considerably elongated in fruit; rachis glabrous or puberulent, trichomes straight or curved, cylindrical. Fruiting pedicels ascending to horizontal, straight, (terete), 5–10(–15) × 0.2–0.3 mm, glabrous or sparsely puberulent adaxially. Flowers: sepals oblong to ovate, 1.5–2.5 × 0.7–1.2 mm; petals white, obovate, (2.5–)3–4(–4.5) × (1–)1.3–2(–2.2) mm, claw 1–1.7 mm; stamens 6; filaments 2–3 mm, (glabrous); anthers 0.4–0.5 mm. Fruits (indehiscent), cordate to subreniform, (2–)2.5–3.7(–4.3) × (3.2–)3.7–5(–5.6) mm, apically (obtuse to subacute), not winged, apical notch absent; valves thin, smooth, reticulate-veined, glabrous; style (0.6–)1–1.8(–2) mm. Seeds ovate, 1.5–2.3 × 1–1.3 mm. 2n = 32, 64.
Phenology: Flowering Apr–Aug.
Habitat: Mountain slopes, roadsides, fields, agricultural lands, stream sides, disturbed grounds, pastures, waste areas
Elevation: 0-3300 m
Distribution
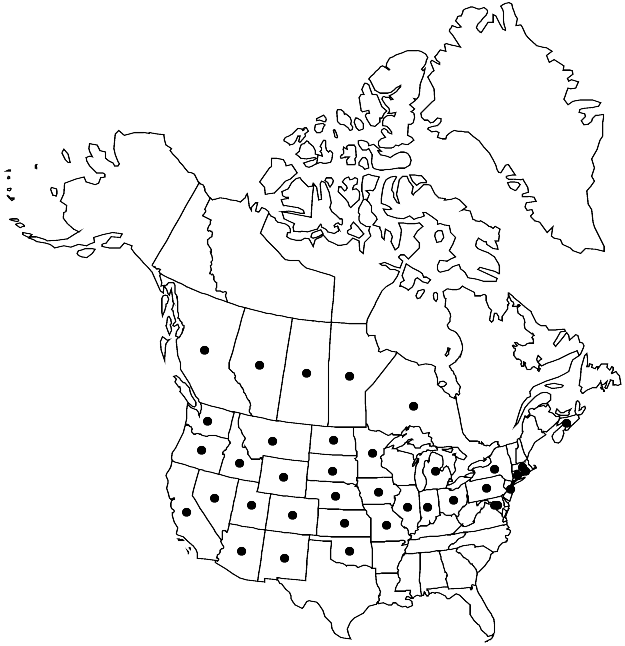
Introduced; Alta., B.C., Man., N.S., Ont., Sask., Ariz., Calif., Colo., Conn., D.C., Idaho, Ill., Ind., Iowa, Kans., Md., Mass., Mich., Minn., Mo., Mont., Nebr., Nev., N.J., N.Mex., N.Y., N.Dak., Ohio, Okla., Oreg., Pa., R.I., S.Dak., Utah, Wash., Wyo., s Europe, sw Asia, introduced also in Mexico (Distrito Federal), South America, s Africa, Australia.
Discussion
Although Lepidium draba is poorly established and known from old collections in the eastern part of the United States, it has become a noxious weed in several western states.
Lepidium draba and its nearest relatives, L. appelianum and L. chalepense, form a monophyletic clade most closely related to L. campestre (K. Mummenhoff et al. 2001). A. Thellung (1906) and C. L. Hitchcock (1936) correctly placed L. draba in Lepidium, as did Linnaeus. The recognition of the first three species in Cardaria and the maintenance of their nearest relative, L. campestre, in Lepidium do not make any sense on both phylogenetic and taxonomic grounds.
Selected References
None.