Spiranthes romanzoffiana
Linnaea 3: 32. 1828.

Plants 8–55 cm. Roots few–several, spreading to descending, tuberous, mostly to 1 cm diam. Leaves persisting through anthesis or fugaceous, basal, often on proximal portion of stem, ascending to spreading, linear to linear-lanceolate, elliptic, or oblanceolate, to 26 × 3 cm. Spikes usually very tightly spiraled, 3 flowers per cycle of spiral, rarely loosely spiraled or with more than 5 flowers per cycle; rachis glabrous to very sparsely pubescent, capitate glands short-stalked or often sessile (longest trichomes 0.03–0.18 mm). Flowers ascending, white to ivory, rarely yellowish white, tubular; sepals and petals connivent, forming hood above abruptly reflexed lip; sepals connate at base, 5.3–12.5 mm; petals linear to ovate, apex obtuse to subacute; lip pandurate, 4.8–10.2 × 1.6–6.8 mm, base broadly expanded, apex broadly dilated, separated by narrow isthmus, apical dilation variable, rarely lip nearly ovate, glabrous or with a few scattered trichomes on adaxial surface; veins typically 3, branches wide-spreading; viscidia linear-lanceolate; ovary 2–7 mm. Seeds monoembryonic. 2n = 44, 66, 88.
Phenology: Flowering Jun–Sep(–Oct).
Habitat: Moist to wet meadows, tundra, marshes, fens, prairies, stream banks, seeps, coastal bluffs, dunes
Elevation: 0–3400 m
Distribution
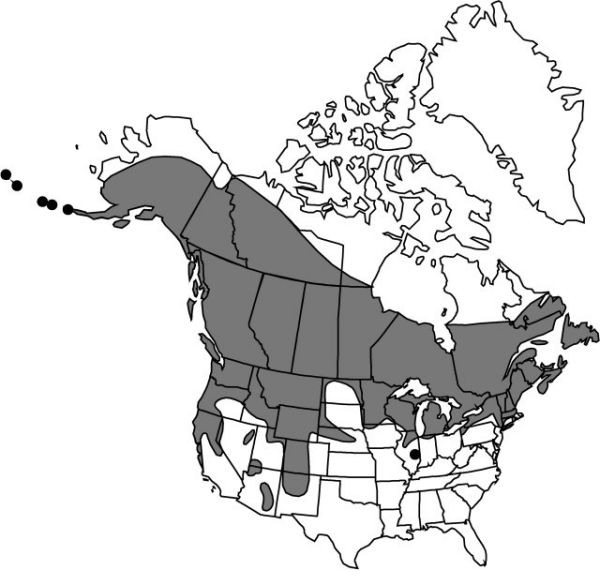
Alta., B.C., Man., N.B., Nfld. and Labr. (Nfld.), N.W.T., N.S., Nunavut, Ont., P.E.I., Que., Sask., Yukon, Alaska, Ariz., Calif., Colo., Conn., Idaho, Ill., Ind., Iowa, Maine, Mass., Mich., Minn., Mont., Nebr., Nev., N.H., N.Mex., N.Y., N.Dak., Oreg., Pa., S.Dak., Utah, Vt., Wash., Wis., Wyo., Europe (Great Britain, n Ireland).
Discussion
Plants of Spiranthes romanzoffiana vary considerably in habit but are usually quite consistent in floral morphology. The strongly hooded, ascending flowers with abruptly reflexed lips provide a distinctive geometric precision. The pandurate lip with typically three veins, the lateral with abruptly wide-spreading branches, is a key feature. In some areas, however, especially at the edges of the range of the species, some variation is apparent and is sometimes coincident with variability in ploidy level. In particular, in California and adjacent southwestern Oregon variability reaches its peak, with plants variously exhibiting yellowish flowers, loosely spiraled inflorescences, and spreading lateral sepals. Some of this variation may result from gene flow from S. porrifolia, but with various ploidy levels common in this area, even within populations, the situation is apparently more complex than simple hybridization.
Hybrids of Spiranthes romanzoffiana with S. lacera var. lacera are known as S. ×simpsonii Catling & Sheviak.
Selected References
None.